
 PDF下载 ( 734 KB)
PDF下载 ( 734 KB)

半乳糖凝集素-9(Gal-9)介导的免疫调节作用在肝脏疾病中的研究进展
DOI: 10.12449/JCH240132
利益冲突声明:本文不存在任何利益冲突。
作者贡献声明:凌珺负责查找文献、撰写论文;朱冰负责拟定写作思路;陈威巍、王卓然负责修改论文;余丽梅、游绍莉负责拟定写作思路,指导撰写文章,修改论文并最后定稿。余丽梅与游绍莉对本文贡献等同,同为第一作者。
Research advances in the immunomodulatory mechanism of galectin-9 and its role in liver diseases
-
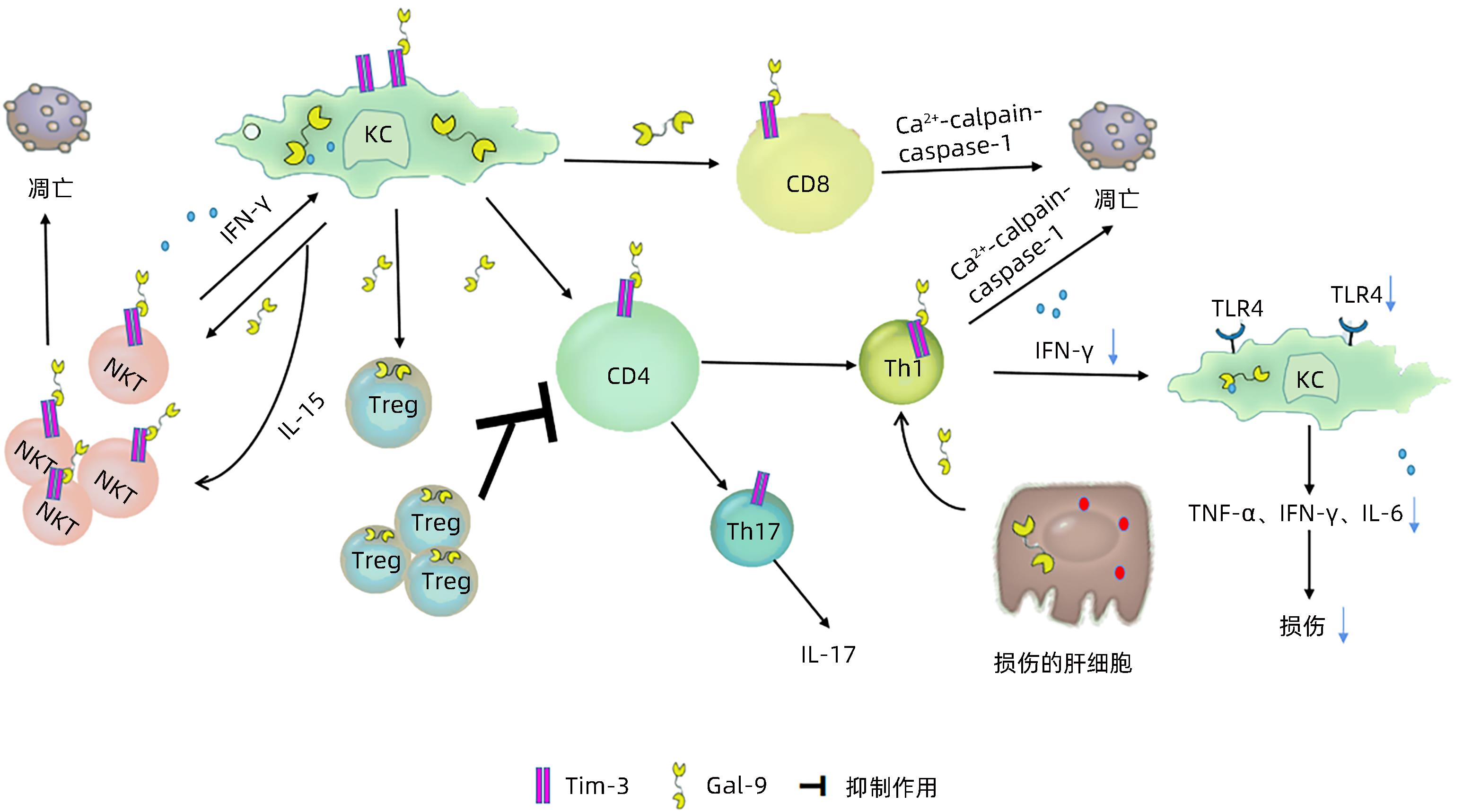
摘要: 半乳糖凝集素-9(Gal-9)是能够特异性识别、结合半乳糖苷的半乳糖凝集素家族成员之一。近年研究发现,Gal-9在肝脏中高水平表达,有助于维持肝内免疫稳态并在多种肝脏疾病中发挥生物学功能。本文综述了Gal-9的免疫调节功能及其在不同肝脏疾病中的作用。Gal-9在不同肝脏疾病中通过多种途径发挥重要的生物学功能,基于对Gal-9具体免疫调节机制和功能的研究,有助于发现Gal-9在肝脏疾病中的治疗作用。Abstract: Galectin-9 (Gal-9) is a member of the galectin family that can specifically recognize and bind to galactosides. Recent studies have shown that Gal-9 is highly expressed in the liver and can help to maintain intrahepatic immune homeostasis and perform biological functions in various liver diseases. This article reviews the immunomodulatory functions of Gal-9 and its role in different liver diseases. Studies have shown that Gal-9 has important biological functions in different liver diseases through multiple pathways. Research on the specific immunomodulatory mechanisms and functions of Gal-9 may help to discover the therapeutic role of Gal-9 in liver diseases.
-
Key words:
- Galectin-9 /
- Immunomodulation /
- Liver Diseases
-
[1] TÜRECI O, SCHMITT H, FADLE N, et al. Molecular definition of a novel human galectin which is immunogenic in patients with Hodgkin’s disease[J]. J Biol Chem, 1997, 272( 10): 6416- 6422. DOI: 10.1074/jbc.272.10.6416. [2] LIU D, ZHU HT, LI CZ. Galectins and galectin-mediated autophagy regulation: New insights into targeted cancer therapy[J]. Biomark Res, 2023, 11( 1): 22. DOI: 10.1186/s40364-023-00466-9. [3] BI SG, EARL LA, JACOBS L, et al. Structural features of galectin-9 and galectin-1 that determine distinct T cell death pathways[J]. J Biol Chem, 2008, 283( 18): 12248- 12258. DOI: 10.1074/jbc.M800523200. [4] JIAO JH, JIAO D, YANG F, et al. Galectin-9 expression predicts poor prognosis in hepatitis B virus-associated hepatocellular carcinoma[J]. Aging, 2022, 14( 4): 1879- 1890. DOI: 10.18632/aging.203909. [5] MOAR P, TANDON R. Galectin-9 as a biomarker of disease severity[J]. Cell Immunol, 2021, 361: 104287. DOI: 10.1016/j.cellimm.2021.104287. [6] MONNEY L, SABATOS CA, GAGLIA JL, et al. Th1-specific cell surface protein Tim-3 regulates macrophage activation and severity of an autoimmune disease[J]. Nature, 2002, 415( 6871): 536- 541. DOI: 10.1038/415536a. [7] LI YR. Mechanisms of gal-9/tim-3 mediated immune regulation[D]. Beijing: Academy of Military Medical Sciences, 2011.李育蓉. Gal-9/Tim-3通路介导免疫调节的机制研究[D]. 北京: 中国人民解放军军事医学科学院, 2011. [8] KANDEL S, ADHIKARY P, LI GF, et al. The TIM3/Gal9 signaling pathway: An emerging target for cancer immunotherapy[J]. Cancer Lett, 2021, 510: 67- 78. DOI: 10.1016/j.canlet.2021.04.011. [9] HARWOOD NMK, GOLDEN-MASON L, CHENG LL, et al. HCV-infected cells and differentiation increase monocyte immunoregulatory galectin-9 production[J]. J Leukoc Biol, 2016, 99( 3): 495- 503. DOI: 10.1189/jlb.5A1214-582R. [10] NACE GW, HUANG H, KLUNE JR, et al. Cellular-specific role of toll-like receptor 4 in hepatic ischemia-reperfusion injury in mice[J]. Hepatology, 2013, 58( 1): 374- 387. DOI: 10.1002/hep.26346. [11] WANG F, XU J, LIAO YL, et al. Tim-3 ligand galectin-9 reduces IL-17 level and accelerates Klebsiella pneumoniae infection[J]. Cell Immunol, 2011, 269( 1): 22- 28. DOI: 10.1016/j.cellimm.2011.03.005. [12] ZHIVOTOVSKY B, BURGESS DH, VANAGS DM, et al. Involvement of cellular proteolytic machinery in apoptosis[J]. Biochem Biophys Res Commun, 1997, 230( 3): 481- 488. DOI: 10.1006/bbrc.1996.6016. [13] KASHIO Y, NAKAMURA K, ABEDIN MJ, et al. Galectin-9 induces apoptosis through the calcium-calpain-caspase-1 pathway[J]. J Immunol, 2003, 170( 7): 3631- 3636. DOI: 10.4049/jimmunol.170.7.3631. [14] EGGENHUIZEN PJ, NG BH, OOI JD. Treg enhancing therapies to treat autoimmune diseases[J]. Int J Mol Sci, 2020, 21( 19): 7015. DOI: 10.3390/ijms21197015. [15] WANG F, WAN LG, ZHANG CC, et al. Tim-3-Galectin-9 pathway involves the suppression induced by CD4+CD25+ regulatory T cells[J]. Immunobiology, 2009, 214( 5): 342- 349. DOI: 10.1016/j.imbio.2008.10.007. [16] MENGSHOL JA, GOLDEN-MASON L, ARIKAWA T, et al. A crucial role for Kupffer cell-derived galectin-9 in regulation of T cell immunity in hepatitis C infection[J]. PLoS One, 2010, 5( 3): e9504. DOI: 10.1371/journal.pone.0009504. [17] SEKI M, OOMIZU S, SAKATA KM, et al. Galectin-9 suppresses the generation of Th17, promotes the induction of regulatory T cells, and regulates experimental autoimmune arthritis[J]. Clin Immunol, 2008, 127( 1): 78- 88. DOI: 10.1016/j.clim.2008.01.006. [18] LV K, ZHANG YY, ZHANG MY, et al. Galectin-9 ameliorates Con A-induced hepatitis by inducing CD4(+)CD25(low/int) effector T-cell apoptosis and increasing regulatory T cell number[J]. PLoS One, 2012, 7( 10): e48379. DOI: 10.1371/journal.pone.0048379. [19] DONG Y, YANG CP, PAN F. Post-translational regulations of Foxp3 in treg cells and their therapeutic applications[J]. Front Immunol, 2021, 12: 626172. DOI: 10.3389/fimmu.2021.626172. [20] MO R, FENG XX, WU YN, et al. Hepatocytes paradoxically affect intrahepatic IFN-γ production in autoimmune hepatitis due to Gal-9 expression and TLR2/4 ligand release[J]. Mol Immunol, 2020, 123: 106- 115. DOI: 10.1016/j.molimm.2020.05.014. [21] RANSON T, VOSSHENRICH CAJ, CORCUFF E, et al. IL-15 availability conditions homeostasis of peripheral natural killer T cells[J]. Proc Natl Acad Sci USA, 2003, 100( 5): 2663- 2668. DOI: 10.1073/pnas.0535482100. [22] TANG ZH, LIANG SW, POTTER J, et al. Tim-3/galectin-9 regulate the homeostasis of hepatic NKT cells in a murine model of nonalcoholic fatty liver disease[J]. J Immunol, 2013, 190( 4): 1788- 1796. DOI: 10.4049/jimmunol.1202814. [23] ZENG XJ. Expression and clinical significance of tim-3/galectin-9 signaling pathway in acute leukemia[D]. Urumqi: Xinjiang Medical University, 2021.曾雪娇. Tim-3/Galectin-9信号通路在急性白血病上的表达及临床意义[D]. 乌鲁木齐: 新疆医科大学, 2021. [24] YANG Y, DONG D, XUE QW, et al. Correlation of negative molecules Tim-3 and Galectin-9 with clinicopathological characteristics of cervical cancer[J]. J Xinjiang Med Univ, 2020, 43( 8): 1004- 1008. DOI: 10.3969/j.issn.1009-5551.2020.08.003.杨艳, 董頔, 薛茜文, 等. 负性分子Tim-3和Galectin-9与宫颈癌患者临床病理特征的相关性研究[J]. 新疆医科大学学报, 2020, 43( 8): 1004- 1008. DOI: 10.3969/j.issn.1009-5551.2020.08.003. [25] COLOMB F, GIRON LB, PREMEAUX TA, et al. Galectin-9 mediates HIV transcription by inducing TCR-dependent ERK signaling[J]. Front Immunol, 2019, 10: 267. DOI: 10.3389/fimmu.2019.00267. [26] MANSOUR AA, RAUCCI F, SAVIANO A, et al. Galectin-9 regulates monosodium urate crystal-induced gouty inflammation through the modulation of treg/Th17 ratio[J]. Front Immunol, 2021, 12: 762016. DOI: 10.3389/fimmu.2021.762016. [27] UCHIDA Y, KE BB, FREITAS MCS, et al. T-cell immunoglobulin mucin-3 determines severity of liver ischemia/reperfusion injury in mice in a TLR4-dependent manner[J]. Gastroenterology, 2010, 139( 6): 2195- 2206. DOI: 10.1053/j.gastro.2010.07.003. [28] TSUNG A, SAHAI R, TANAKA H, et al. The nuclear factor HMGB1 mediates hepatic injury after murine liver ischemia-reperfusion[J]. J Exp Med, 2005, 201( 7): 1135- 1143. DOI: 10.1084/jem.20042614. [29] ZHAI Y, SHEN XD, O’CONNELL R, et al. Cutting edge: TLR4 activation mediates liver ischemia/reperfusion inflammatory response via IFN regulatory factor 3-dependent MyD88-independent pathway[J]. J Immunol, 2004, 173( 12): 7115- 7119. DOI: 10.4049/jimmunol.173.12.7115. [30] ZHAO LZ, LIANG J, RAO W, et al. Cross-regulation by TLR4 and T cell Ig mucin-3 determines severity of liver injury in a CCl4-induced mouse model[J]. Scand J Immunol, 2020, 91( 4): e12851. DOI: 10.1111/sji.12851. [31] LIU YX, JI HF, ZHANG Y, et al. Recipient T cell TIM-3 and hepatocyte galectin-9 signalling protects mouse liver transplants against ischemia-reperfusion injury[J]. J Hepatol, 2015, 62( 3): 563- 572. DOI: 10.1016/j.jhep.2014.10.034. [32] HASSAN A, FONTANA RJ. The diagnosis and management of idiosyncratic drug-induced liver injury[J]. Liver Int, 2019, 39( 1): 31- 41. DOI: 10.1111/liv.13931. [33] ROSEN HR, BIGGINS SW, NIKI T, et al. Association between plasma level of galectin-9 and survival of patients with drug-induced acute liver failure[J]. Clin Gastroenterol Hepatol, 2016, 14( 4): 606- 612.e3. DOI: 10.1016/j.cgh.2015.09.040. [34] ROSEN HR. Emerging concepts in immunity to hepatitis C virus infection[J]. J Clin Invest, 2013, 123( 10): 4121- 4130. DOI: 10.1172/JCI67714. [35] YANG XF, WANG LX, HUANG CX, et al. Mechanism of gamma-chain cytokines in regulating the expression of T-cell immunoglobulin and mucin domain-containing molecule 3 in CD8+ T cells of chronic hepatitis B patients[J]. J Clin Hepatol, 2021, 37( 5): 1059- 1064. DOI: 10.3969/j.issn.1001-5256.2021.05.017.杨晓飞, 王临旭, 黄长形, 等. γ链细胞因子对慢性乙型肝炎患者T淋巴细胞免疫球蛋白黏蛋白分子3表达调控的诱导机制[J]. 临床肝胆病杂志, 2021, 37( 5): 1059- 1064. DOI: 10.3969/j.issn.1001-5256.2021.05.017. [36] NEBBIA G, PEPPA D, SCHURICH A, et al. Upregulation of the Tim-3/galectin-9 pathway of T cell exhaustion in chronic hepatitis B virus infection[J]. PLoS One, 2012, 7( 10): e47648. DOI: 10.1371/journal.pone.0047648. [37] GOLDEN-MASON L, PALMER BE, KASSAM N, et al. Negative immune regulator Tim-3 is overexpressed on T cells in hepatitis C virus infection and its blockade rescues dysfunctional CD4+ and CD8+ T cells[J]. J Virol, 2009, 83( 18): 9122- 9130. DOI: 10.1128/JVI.00639-09. [38] McMAHAN RH, GOLDEN-MASON L, NISHIMURA MI, et al. Tim-3 expression on PD-1+ HCV-specific human CTLs is associated with viral persistence, and its blockade restores hepatocyte-directed in vitro cytotoxicity[J]. J Clin Invest, 2010, 120( 12): 4546- 57. DOI: 10.1172/JCI43127. [39] LAI JH, LUO SF, WANG MY, et al. Translational implication of galectin-9 in the pathogenesis and treatment of viral infection[J]. Int J Mol Sci, 2017, 18( 10): 2108. DOI: 10.3390/ijms18102108. [40] LIU SY, XU C, YANG F, et al. Natural killer cells induce CD8+ T cell dysfunction via galectin-9/TIM-3 in chronic hepatitis B virus infection[J]. Front Immunol, 2022, 13: 884290. DOI: 10.3389/fimmu.2022.884290. [41] LIBERAL R, GRANT CR, HOLDER BS, et al. The impaired immune regulation of autoimmune hepatitis is linked to a defective galectin-9/tim-3 pathway[J]. Hepatology, 2012, 56( 2): 677- 686. DOI: 10.1002/hep.25682. [42] JIANG Y, HAN QJ, ZHAO HJ, et al. The mechanisms of HBV-induced hepatocellular carcinoma[J]. J Hepatocell Carcinoma, 2021, 8: 435- 450. DOI: 10.2147/JHC.S307962. [43] LI H, WU K, TAO KX, et al. Tim-3/galectin-9 signaling pathway mediates T-cell dysfunction and predicts poor prognosis in patients with hepatitis B virus-associated hepatocellular carcinoma[J]. Hepatology, 2012, 56( 4): 1342- 1351. DOI: 10.1002/hep.25777. [44] GOLDEN-MASON L, MCMAHAN RH, STRONG M, et al. Galectin-9 functionally impairs natural killer cells in humans and mice[J]. J Virol, 2013, 87( 9): 4835- 4845. DOI: 10.1128/JVI.01085-12. [45] FUJITA K, IWAMA H, SAKAMOTO T, et al. Galectin-9 suppresses the growth of hepatocellular carcinoma via apoptosis in vitro and in vivo[J]. Int J Oncol, 2015, 46( 6): 2419- 2430. DOI: 10.3892/ijo.2015.2941. -

 下载:
下载:
 本文二维码
本文二维码
图(1)
计量
- 文章访问数: 1730
- HTML全文浏览量: 638
- PDF下载量: 103
- 被引次数: 0

