
 PDF下载 ( 920 KB)
PDF下载 ( 920 KB)

中国旱獭Ⅰ型干扰素受体β亚基克隆、表达及功能初步鉴定
DOI: 10.12449/JCH240210
Preliminary identification of the cloning, expression, and function of Marmota himalayana type I interferon receptor β subunit
-
摘要:
目的 克隆中国旱獭Ⅰ型干扰素受体β亚基(mhIFNAR2)的基因,并进行抗体制备及功能鉴定。 方法 应用RT-PCR技术,从中国旱獭脾组织中扩增得到序列,克隆至原核表达载体pRSET-B,表达重组蛋白,电泳和Western Blot法鉴定;重组蛋白常规免疫BALB/c小鼠制备其胞外段多克隆抗体,免疫组化、免疫荧光和Western Blot法鉴定;再通过siRNA阻断的方法检测其功能。计量资料多组间比较采用方差分析,进一步两两比较采用LSD-t检验。 结果 从mhIFNAR2扩增出149~1 300 bp片段,其同源性在分析的种属中以土拨鼠最高,可达98.05%。成功地构建了表达胞外段mhIFNAR2(50-181aa)蛋白的原核表达质粒,命名为pRSET-B.mhIFNAR2;其表达重组蛋白分子量27 kD,纯化后纯度约为95%,浓度约为160 μg/mL。用纯化的重组蛋白常规免疫BALB/c小鼠后,获得1∶1 000的特异性多克隆抗体,用免疫组化及免疫荧光可见细胞膜、细胞质有表达。合成的三条siRNA,其中有一条起始于277位点的siRNA(siRNA277)与空白对照及阴性对照相比,可以沉默目的基因的表达,并能减弱干扰素的信号通路(P值均<0.05)。 结论 获得mhIFNAR2的部分序列,成功地制备出抗mhIFNAR2胞外段多克隆抗体,该抗体有较高的效价和特异性,并能用于免疫组化、免疫荧光及Western Blot的检测。用siRNA277可以抑制目的基因的表达,并能阻断干扰素的信号通路。 Abstract:Objective To clone the gene of Marmota himalayana type Ⅰ interferon receptor β subunit (mhIFNAR2), and to perform antibody preparation and functional identification. Methods RT-PCR was used for amplification in the spleen tissue of Marmota himalayana to obtain the sequence, which was cloned to the prokaryotic expression vector pRSET-B to express the recombinant protein. Electrophoresis and Western blot were used for identification. BALB/c mice were immunized with the recombinant protein to prepare the polyclonal antibody of its extracellular domain; immunohistochemistry, immunofluorescence assay, and Western Blot were used for identification, and the method of siRNA blockade was used to investigate its function. An analysis of variance was used for comparison of continuous data between multiple groups, and the least significant difference t-test was used for comparison between two groups. Results A fragment of mhIFNAR2 (149 — 1 300 bp) was obtained from spleen tissue, which showed the highest homology of 98.05% in marmot. A prokaryotic expression plasmid was successfully constructed for expression of the extracellular domain of the mhIFNAR2(50-181aa) and was named pRSET-B.mhIFNAR2, and the recombinant protein expressed by this plasmid had a molecular weight of 27 kD, a purity of about 95% after purification, and a concentration of 160 μg/mL. After BALB/c mice were immunized with the purified recombinant protein, 1∶1 000 specific polyclonal antibodies were obtained, and immunohistochemistry and immunofluorescence assay showed the expression in cell membrane and cytoplasm. Among the three siRNAs synthesized, the siRNA starting from the 277 locus (siRNA277) could silence the expression of target genes and weaken the interferon signaling pathway compared with the blank control group and the negative control group (both P<0.05). Conclusion The fragment of mhIFNAR2 is obtained, and the polyclonal antibody for the extracellular domain of mhIFNAR2 is successfully prepared, with relatively high titer and specificity, and can be used for immunohistochemistry, immunofluorescence assay, and Western blot. -
Key words:
- Receptor, Interferon alpha-beta /
- Clone /
- Hepatitis B
-
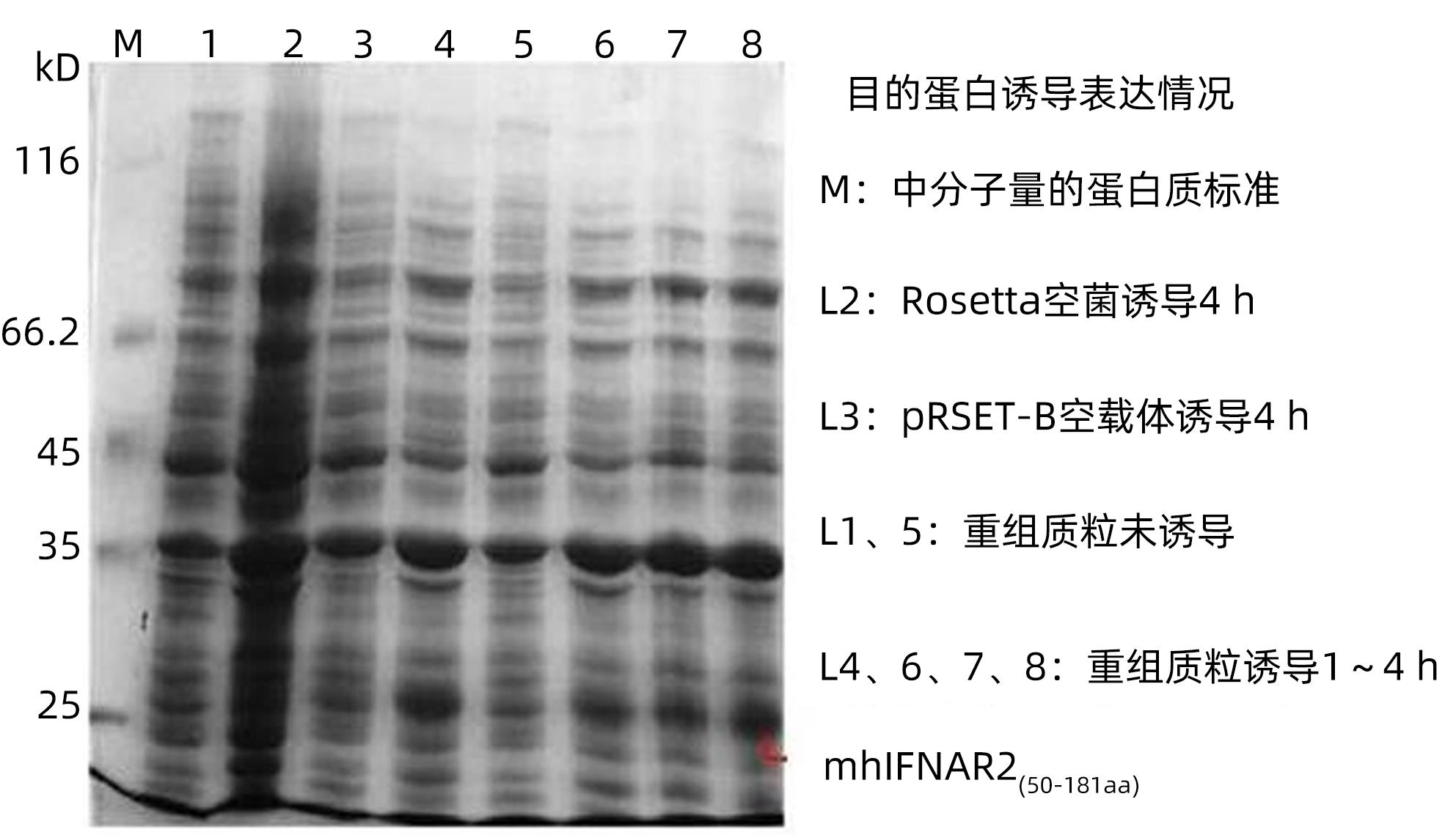
图 2 SDS-PAGE电泳分析His-mhIFNAR2蛋白的表达
Figure 2. Expression of His-mhIFNAR2 analyzed by SDS-PAGE
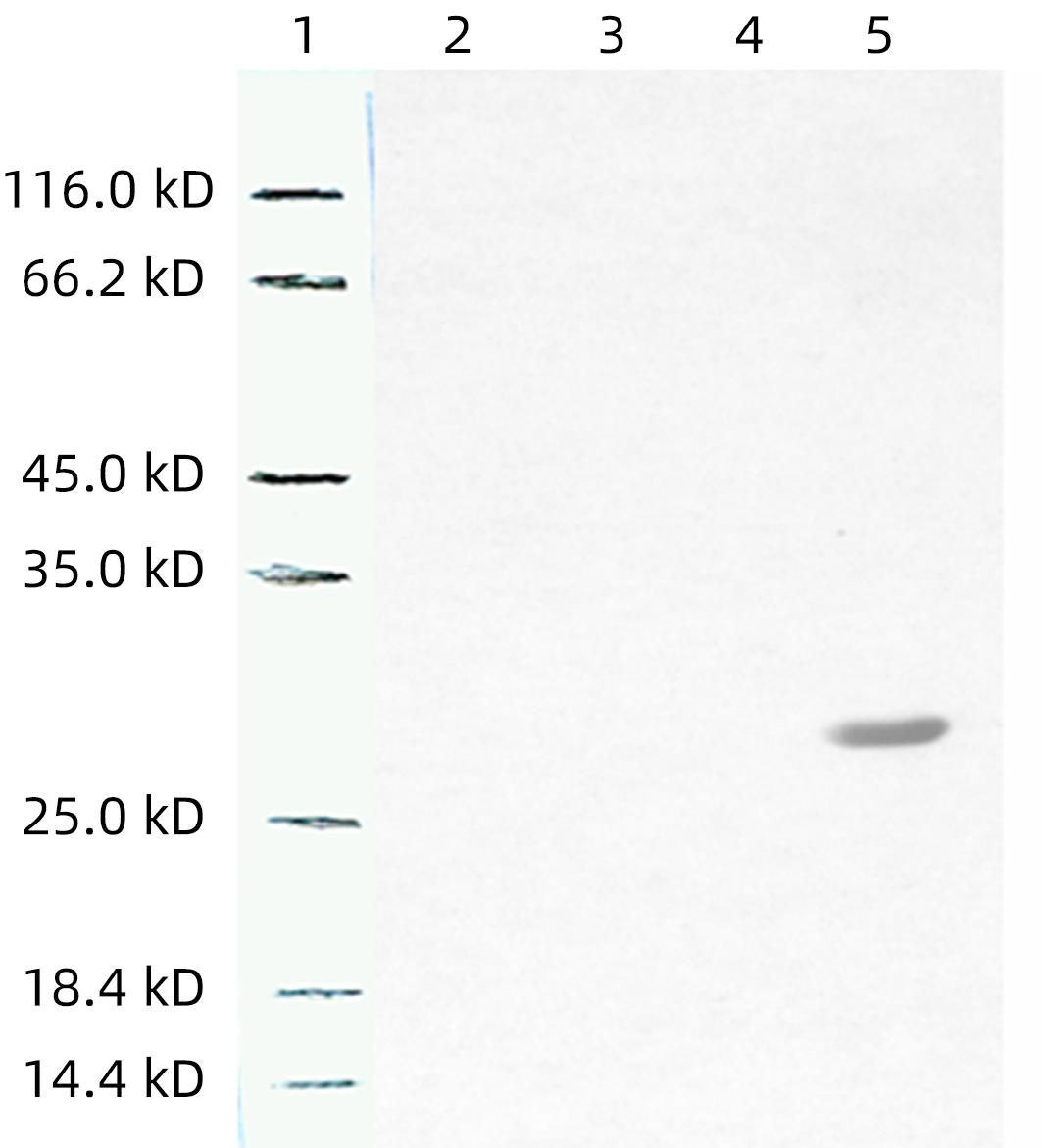
注: L1,中分子量的蛋白质标准;L2,mhIFNAR2多抗血清作为一抗(1∶1 000);L3,mhIFNAR2多抗血清作为一抗(1∶10 000);L4,正常小鼠血清作为一抗;L5,兔抗His作为一抗。
图 4 纯化的目的蛋白Western Blot检测
Figure 4. Western Blot analysis of the activity and specificity of antisera

图 5 mhIFNAR2多克隆抗体的免疫组化结果(DAB,×400)
Figure 5. Immunohistochemical of mhIFNAR2 polyclonal antibodies(DAB,×400)
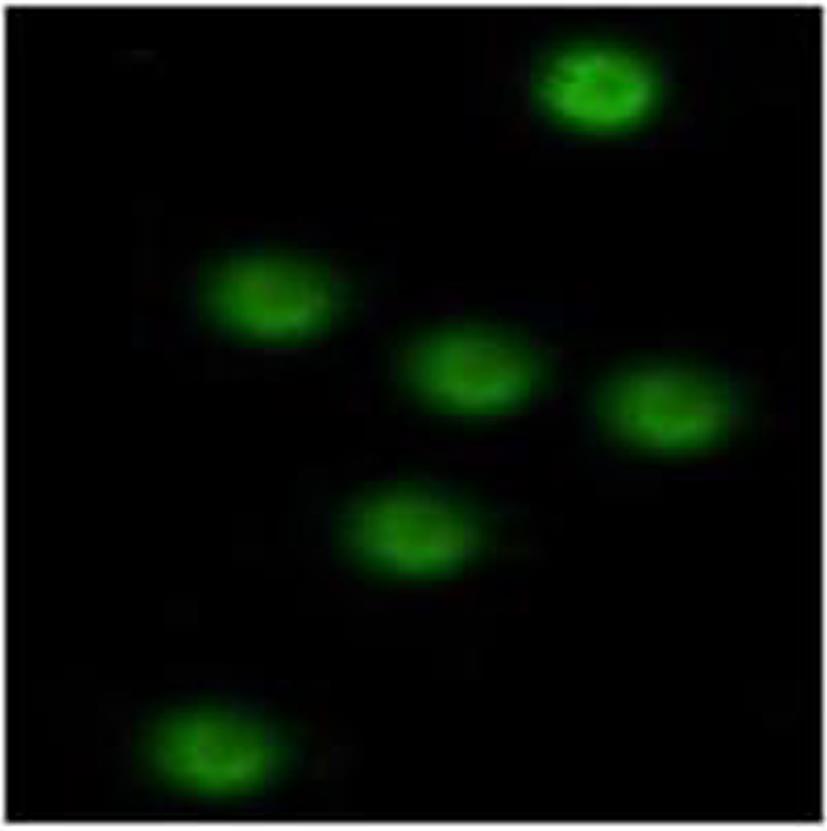
图 6 PBMC中mhIFNAR2免疫荧光检测
Figure 6. Stained with mouse anti-mhIFNAR2 antibody by immunofluorescence in PBMC

图 7 不同浓度的siRNA840对WH12-6细胞mhIFNAR2、M×A基因的抑制作用
Figure 7. The inhibitory effect of siRNA840 at different concentrations on mhIFNAR2 and M×A
表 1 干扰素受体β亚基的引物序列
Table 1. Primer sequences for interferon receptor β subunit
名称 序列 核苷酸 位点 mhIFNAR1-2s 5′-AGTACATTTAGAAGCTGAAG-3' 396~416 mhIFNAR1-2as 5'-CTCTTCAGACCAAAAAGATG-3' 943~963 mhIFNAR2-1s 5'-CCATCTTATCATGGGAATTA-3' 149~169 mhIFNAR2-1as 5'-CTCTCAAAAACACAGAGTT-3' 1 281~1 300  下载: 导出CSV
下载: 导出CSV
表 2 mhIFNAR2 siRNA 模板序列
Table 2. Template of mhIFNAR2 siRNA
cDNA 正义链(5′-3′) 反义链(3′-5′) GAAGTAGCTCTCAGAACTA(mhIFNAR1B840B) GAAGUAGCUCUCAGAACUAdTdT dTdTCUUCAUCGAGAGUCUUGAU GCTTTACAGACCACATTAA(mhIFNAR2B277B) GCUUUACAGACCACAUUAAdTdT dTdTCGAAAUGUCUGGUGUAAUU GCTGAAGATAAGGCAATAA(mhIFNAR1B412B) GCUGAAGAUAAGGCAAUAAdTdT dTdTCGACUUCUAUUCCGUUAUU 阴性对照 公司未提供序列
下载: 导出CSV
表 3 EMCV感染滴度测定结果
Table 3. Result of EMCV infection titer determination
病毒稀释 有CPE孔 无CPE孔 累计有CPE孔 累计无CPE孔 有/总和 感染百分率(%) 100 4 0 39 0 39/39 100 10-1 4 0 35 0 35/35 100 10-2 4 0 31 0 31/31 100 10-3 4 0 27 0 27/27 100 10-4 4 0 23 0 23/23 100 10-5 4 0 19 0 19/19 100 10-6 4 0 15 0 15/15 100 10-7 4 0 11 0 11/11 100 10-8 4 0 7 0 7/7 100 10-9 2 2 3 2 3/5 60 10-10 1 3 1 5 1/6 16.7
下载: 导出CSV
表 4 不同浓度的siRNA277对WH12-6细胞mhIFNAR2、M×A基因的抑制作用
Table 4. The inhibitory effect of siRNA277 at different concentrations on mhIFNAR2 and M×A
组别 mhIFNAR2 M×A 空白对照(n=3) 11.678±0.725 3.087±1.910 阴性对照(n=3) 14.190±3.100 4.150±0.500 20 nmol/L(n=3) 2.538±0.1241)2) 1.350±0.2121)2) 注:与空白对照组比较,1)P<0.05;与阴性对照组比较,(2)P<0.05。
下载: 导出CSV
-
[1] ERSVAER E, SKAVLAND J, ULVESTAD E, et al. Effects of interferon gamma on native human acute myelogenous leukaemia cells[J]. Cancer Immunol Immunother, 2007, 56( 1): 13- 24. DOI: 10.1007/s00262-006-0159-1. [2] MERLI P, QUINTARELLI C, STROCCHIO L, et al. The role of interferon-gamma and its signaling pathway in pediatric hematological disorders[J]. Pediatr Blood Cancer, 2021, 68( 4): e28900. DOI: 10.1002/pbc.28900. [3] SCHRODER K, HERTZOG PJ, RAVASI T, et al. Interferon-gamma: An overview of signals, mechanisms and functions[J]. J Leukoc Biol, 2004, 75( 2): 163- 189. DOI: 10.1189/jlb.0603252. [4] BHAT MY, SOLANKI HS, ADVANI J, et al. Comprehensive network map of interferon gamma signaling[J]. J Cell Commun Signal, 2018, 12( 4): 745- 751. DOI: 10.1007/s12079-018-0486-y. [5] WANG BJ, TIAN YJ, MENG ZJ, et al. Establishing a new animal model for hepadnaviral infection: Susceptibility of Chinese Marmota-species to woodchuck hepatitis virus infection[J]. J Gen Virol, 2011, 92( Pt 3): 681- 691. DOI: 10.1099/vir.0.025023-0. [6] GUO WN, ZHU B, AI L, et al. Animal models for the study of hepatitis B virus infection[J]. Zool Res, 2018, 39( 1): 25- 31. DOI: 10.24272/j.issn.2095-8137.2018.013. [7] FAN HB, ZHU ZN, WANG Y, et al. Molecular characterization of the type I IFN receptor in two woodchuck species and detection of its expression in liver samples from woodchucks infected with woodchuck hepatitis virus(WHV)[J]. Cytokine, 2012, 60( 1): 179- 185. DOI: 10.1016/j.cyto.2012.05.013.2012. [8] SCHREIBER G. The role of type I interferons in the pathogenesis and treatment of COVID-19[J]. Front Immunol, 2020, 11: 595739. DOI: 10.3389/fimmu.2020.595739. [9] MCNAB F, MAYER-BARBER K, SHER A, et al. Type I interferons in infectious disease[J]. Nat Rev Immunol, 2015, 15( 2): 87- 103. DOI: 10.1038/nri3787. [10] MURIRA A, LAMARRE A. Type-I interferon responses: From friend to foe in the battle against chronic viral infection[J]. Front Immunol, 2016, 7: 609. DOI: 10.3389/fimmu.2016.00609. [11] BASTARD P, MANRY J, CHEN J, et al. Herpes simplex encephalitis in a patient with a distinctive form of inherited IFNAR1 deficiency[J]. J Clin Invest, 2021, 131( 1): e139980. DOI: 10.1172/JCI139980. [12] KHANMOHAMMADI S, REZAEI N, KHAZAEI M, et al. A case of autosomal recessive interferon alpha/beta receptor alpha chain(IFNAR1) deficiency with severe COVID-19[J]. J Clin Immunol, 2022, 42( 1): 19- 24. DOI: 10.1007/s10875-021-01166-5. [13] PLATANIAS LC. Mechanisms of type-I- and type-II-interferon-mediated signalling[J]. Nat Rev Immunol, 2005, 5( 5): 375- 386. DOI: 10.1038/nri1604. [14] TOUGH DF, SUN S, ZHANG X, et al. Stimulation of naïve and memory T cells by cytokines[J]. Immunol Rev, 1999, 170: 39- 47. DOI: 10.1111/j.1600-065x.1999.tb01327.x. [15] World Health Organization. Global Hepatitis Report 2017[EB/OL].( 2017-04-19)[ 2023-04-11]. https://www.who.int/hepatitis/publications/global-hepatitis-report2017/en/. https://www.who.int/hepatitis/publications/global-hepatitis-report2017 /en/ [16] LU YP, XU Y, YANG DL, et al. Molecular characterization of woodchuck type I interferons and their expression by woodchuck peripheral blood lymphocytes[J]. Cytokine, 2008, 41( 2): 127- 135. DOI: 10.1016/j.cyto.2007.11.002. [17] WANG BJ, LOHRENGEL B, LU YP, et al. Molecular characterization of woodchuck interleukin 15(wIL-15) and detection of its expression in liver samples of woodchucks infected with woodchuck hepatitis virus(WHV)[J]. Cytokine, 2005, 32( 6): 296- 303. DOI: 10.1016/j.cyto.2005.11.007. [18] YANG Y, HUANG H, ZHANG ZH, et al. Cloning, expression and polyclonal antibody preparation of the asialoglycoprotein receptor of Marmota Himalayan[J]. J Huazhong Univ Sci Technol(Med Sci), 2007, 27( 4): 411- 414. DOI: 10.1007/s11596-007-0415-4. [19] SHEEHAN KC, LAI KS, DUNN GP, et al. Blocking monoclonal antibodies specific for mouse IFN-alpha/beta receptor subunit 1(IFNAR-1) from mice immunized by in vivo hydrodynamic transfection[J]. J Interferon Cytokine Res, 2006, 26( 11): 804- 819. DOI: 10.1089/jir.2006.26.804. [20] BENOIT P, MAGUIRE D, PLAVEC I, et al. A monoclonal antibody to recombinant human IFN-alpha receptor inhibits biologic activity of several species of human IFN-alpha, IFN-beta, and IFN-omega. Detection of heterogeneity of the cellular type I IFN receptor[J]. J Immunol, 1993, 150( 3): 707- 716. [21] COLAMONICI OR, D’ALESSANDRO F, DIAZ MO, et al. Characterization of three monoclonal antibodies that recognize the interferon alpha 2 receptor[J]. Proc Natl Acad Sci U S A, 1990, 87( 18): 7230- 7234. DOI: 10.1073/pnas.87.18.7230. [22] UZÉ G, LUTFALLA G, EID P, et al. Murine tumor cells expressing the gene for the human interferon alpha beta receptor elicit antibodies in syngeneic mice to the active form of the receptor[J]. Eur J Immunol, 1991, 21( 2): 447- 451. DOI: 10.1002/eji.1830210229. -

 本文二维码
本文二维码
计量
- 文章访问数: 846
- HTML全文浏览量: 320
- PDF下载量: 60
- 被引次数: 0

