
 PDF下载 ( 649 KB)
PDF下载 ( 649 KB)

滤泡辅助性T淋巴细胞在自身免疫性肝炎中的作用机制
DOI: 10.12449/JCH240727
利益冲突声明:本文不存在任何利益冲突。
作者贡献声明:罗龙龙、王丽菲负责检索文献,撰写论文;邢国静、郑英、卢利霞、李初谊、李斌、马俊参与修改论文;于晓辉、张久聪负责拟定写作思路,指导撰写文章并最后定稿。
-
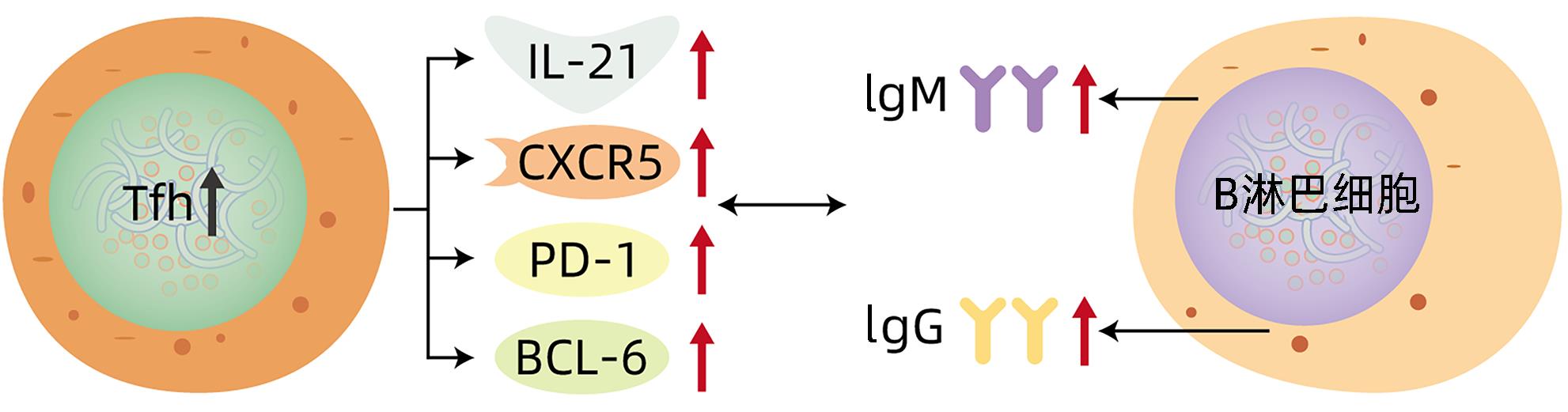
摘要: 自身免疫性肝炎(AIH)是由自身免疫系统攻击肝细胞导致的慢性肝炎,随着病程的延长,可逐渐发展为肝硬化,甚至肝癌。尽管目前对于AIH的认识和治疗已取得很大进展,但其病因和发病机制仍不明确。T淋巴细胞在AIH的发生、发展过程中发挥至关重要的作用,本文聚焦于滤泡辅助性T淋巴细胞,阐述其在AIH机制中的研究进展,以期为AIH的临床治疗提供新的思路和策略。
-
关键词:
- 肝炎, 自身免疫性 /
- 滤泡性辅助性T细胞 /
- T淋巴细胞, 调节性 /
- B-淋巴细胞
Abstract: Autoimmune hepatitis (AIH) is a type of chronic hepatitis caused by the attack of hepatocytes by the autoimmune system, and with the prolongation of disease course, it may gradually progress to liver cirrhosis and even hepatocellular carcinoma. Although great achievements have been made in the understanding and treatment of AIH, its etiology and pathogenesis still remain unclear. T cells play a crucial role in the development and progression of AIH, and by focusing on follicular helper T cells, this article elaborates on the research advances in follicular helper T cells in AIH, in order to provide new ideas and strategies for the clinical treatment of AIH. -
[1] CARDON A, CONCHON S, RENAND A. Mechanisms of autoimmune hepatitis[J]. Curr Opin Gastroenterol, 2021, 37( 2): 79- 85. DOI: 10.1097/MOG.0000000000000704. [2] DALEKOS GN, SAMAKIDOU A, LYBEROPOULOU A, et al. Recent advances in the diagnosis and management of autoimmune hepatitis[J]. Pol Arch Intern Med, 2022, 132( 9): 16334. DOI: 10.20452/pamw.16334. [3] OLIVAS I, RODRÍGUEZ-TAJES S, LONDOÑO MC. Autoimmune hepatitis: Challenges and novelties[J]. Med Clin(Barc), 2022, 159( 6): 289- 298. DOI: 10.1016/j.medcli.2022.04.004. [4] SIRBE C, SIMU GL, SZABO I, et al. Pathogenesis of autoimmune hepatitis-cellular and molecular mechanisms[J]. Int J Mol Sci, 2021, 22( 24): 13578. DOI: 10.3390/ijms222413578. [5] SONG WZ, CRAFT J. T follicular helper cell heterogeneity: Time, space, and function[J]. Immunol Rev, 2019, 288( 1): 85- 96. DOI: 10.1111/imr.12740. [6] SCHROEDER AR, ZHU FM, HU H. Stepwise Tfh cell differentiation revisited: New advances and long-standing questions[J]. Fac Rev, 2021, 10: 3. DOI: 10.12703/r/10-3. [7] NANCE JP, BÉLANGER S, JOHNSTON RJ, et al. Bcl6 middle domain repressor function is required for T follicular helper cell differentiation and utilizes the corepressor MTA3[J]. Proc Natl Acad Sci U S A, 2015, 112( 43): 13324- 13329. DOI: 10.1073/pnas.1507312112. [8] GENSOUS N, CHARRIER M, DULUC D, et al. T follicular helper cells in autoimmune disorders[J]. Front Immunol, 2018, 9: 1637. DOI: 10.3389/fimmu.2018.01637. [9] KHANAM A, AYITHAN N, TANG L, et al. IL-21-deficient T follicular helper cells support B cell responses through IL-27 in patients with chronic hepatitis B[J]. Front Immunol, 2021, 11: 599648. DOI: 10.3389/fimmu.2020.599648. [10] POWELL MD, READ KA, SREEKUMAR BK, et al. IL-12 signaling drives the differentiation and function of a TH1-derived TFH1-like cell population[J]. Sci Rep, 2019, 9( 1): 13991. DOI: 10.1038/s41598-019-50614-1. [11] MESAS-FERNÁNDEZ A, BODNER E, HILKE FJ, et al. Interleukin-21 in autoimmune and inflammatory skin diseases[J]. Eur J Immunol, 2023, 53( 4): e2250075. DOI: 10.1002/eji.202250075. [12] ZANDER R, KASMANI MY, CHEN Y, et al. Tfh-cell-derived interleukin 21 sustains effector CD8+ T cell responses during chronic viral infection[J]. Immunity, 2022, 55( 3): 475- 493. DOI: 10.1016/j.immuni.2022.01.018. [13] SHI JW, HOU SY, FANG Q, et al. PD-1 controls follicular T helper cell positioning and function[J]. Immunity, 2018, 49( 2): 264- 274. DOI: 10.1016/j.immuni.2018.06.012. [14] TANEMURA S, TSUJIMOTO H, SEKI N, et al. Role of interferons(IFNs) in the differentiation of T peripheral helper(Tph) cells[J]. Int Immunol, 2022, 34( 10): 519- 532. DOI: 10.1093/intimm/dxac026. [15] PAPILLION A, POWELL MD, CHISOLM DA, et al. Inhibition of IL-2 responsiveness by IL-6 is required for the generation of GC-TFH cells[J]. Sci Immunol, 2019, 4( 39): eaaw7636. DOI: 10.1126/sciimmunol.aaw7636. [16] CROTTY S. T follicular helper cell biology: A decade of discovery and diseases[J]. Immunity, 2019, 50( 5): 1132- 1148. DOI: 10.1016/j.immuni.2019.04.011. [17] WANG YF, SHI JW, YAN JC, et al. Germinal-center development of memory B cells driven by IL-9 from follicular helper T cells[J]. Nat Immunol, 2017, 18( 8): 921- 930. DOI: 10.1038/ni.3788. [18] PAPA I, SALIBA D, PONZONI M, et al. TFH-derived dopamine accelerates productive synapses in germinal centres[J]. Nature, 2017, 547( 7663): 318- 323. DOI: 10.1038/nature23013. [19] MARSMAN C, VERSTEGEN NJ, STREUTKER M, et al. Termination of CD40L co-stimulation promotes human B cell differentiation into antibody-secreting cells[J]. Eur J Immunol, 2022, 52( 10): 1662- 1675. DOI: 10.1002/eji.202249972. [20] GARDELL JL, PARKER DC. CD40L is transferred to antigen-presenting B cells during delivery of T-cell help[J]. Eur J Immunol, 2017, 47( 1): 41- 50. DOI: 10.1002/eji.201646504. [21] MICHEL NA, ZIRLIK A, WOLF D. CD40L and its receptors in atherothrombosis-an update[J]. Front Cardiovasc Med, 2017, 4: 40. DOI: 10.3389/fcvm.2017.00040. [22] LU J, WU J, XIE FT, et al. CD4+ T cell-released extracellular vesicles potentiate the efficacy of the HBsAg vaccine by enhancing B cell responses[J]. Adv Sci(Weinh), 2019, 6( 23): 1802219. DOI: 10.1002/advs.201802219. [23] FERNÁNDEZ-MESSINA L, RODRÍGUEZ-GALÁN A, DE YÉBENES VG, et al. Transfer of extracellular vesicle-microRNA controls germinal center reaction and antibody production[J]. EMBO Rep, 2020, 21( 4): e48925. DOI: 10.15252/embr.201948925. [24] MA L, QIN JJ, JI HF, et al. Tfh and plasma cells are correlated with hypergammaglobulinaemia in patients with autoimmune hepatitis[J]. Liver Int, 2014, 34( 3): 405- 415. DOI: 10.1111/liv.12245. [25] MORITA R, SCHMITT N, BENTEBIBEL SE, et al. Human blood CXCR5+CD4+ T cells are counterparts of T follicular cells and contain specific subsets that differentially support antibody secretion[J]. Immunity, 2011, 34( 1): 108- 121. DOI: 10.1016/j.immuni.2010.12.012. [26] ABE K, TAKAHASHI A, IMAIZUMI H, et al. Interleukin-21 plays a critical role in the pathogenesis and severity of type I autoimmune hepatitis[J]. Springerplus, 2016, 5( 1): 777. DOI: 10.1186/s40064-016-2512-y. [27] MA L, ZHANG LW, ZHUANG Y, et al. Lactobacillus improves the effects of prednisone on autoimmune hepatitis via gut microbiota-mediated follicular helper T cells[J]. Cell Commun Signal, 2022, 20( 1): 83. DOI: 10.1186/s12964-021-00819-7. [28] WANG WJ, SUNG N, GILMAN-SACHS A, et al. T helper(Th) cell profiles in pregnancy and recurrent pregnancy losses: Th1/Th2/Th9/Th17/Th22/tfh cells[J]. Front Immunol, 2020, 11: 2025. DOI: 10.3389/fimmu.2020.02025. [29] ZHANG R, ZHAO Y, CHEN X, et al. Low-dose IL-2 therapy in autoimmune diseases: An update review[J]. Int Rev Immunol, 2024, 43( 3): 113- 137. DOI: 10.1080/08830185.2023.2274574. [30] KIMURA N, YAMAGIWA S, SUGANO T, et al. Usefulness of chemokine C-C receptor 7-/programmed cell death-1+ follicular helper T cell subset frequencies in the diagnosis of autoimmune hepatitis[J]. Hepatol Res, 2019, 49( 9): 1026- 1033. DOI: 10.1111/hepr.13356. [31] CHENG ZL, YANG L, CHU HK. The gut microbiota: A novel player in autoimmune hepatitis[J]. Front Cell Infect Microbiol, 2022, 12: 947382. DOI: 10.3389/fcimb.2022.947382. [32] DING TT, SU R, WU RH, et al. Frontiers of autoantibodies in autoimmune disorders: Crosstalk between tfh/tfr and regulatory B cells[J]. Front Immunol, 2021, 12: 641013. DOI: 10.3389/fimmu.2021.641013. [33] LINTERMAN MA, PIERSON W, LEE SK, et al. Foxp3+ follicular regulatory T cells control the germinal center response[J]. Nat Med, 2011, 17( 8): 975- 982. DOI: 10.1038/nm.2425. [34] CHUNG Y, TANAKA S, CHU FL, et al. Follicular regulatory T cells expressing Foxp3 and Bcl-6 suppress germinal center reactions[J]. Nat Med, 2011, 17( 8): 983- 988. DOI: 10.1038/nm.2426. [35] MA L, ZHANG LW, DAI J, et al. Dysregulated TFR and TFH cells correlate with B-cell differentiation and antibody production in autoimmune hepatitis[J]. J Cell Mol Med, 2020, 24( 7): 3948- 3957. DOI: 10.1111/jcmm.14997. [36] MA L, ZHANG LW, SONG JG, et al. Fecal microbiota transplantation controls progression of experimental autoimmune hepatitis in mice by modulating the TFR/TFH immune imbalance and intestinal microbiota composition[J]. Front Immunol, 2021, 12: 728723. DOI: 10.3389/fimmu.2021.728723. -

 下载:
下载:
 本文二维码
本文二维码
图(1)
计量
- 文章访问数: 1045
- HTML全文浏览量: 306
- PDF下载量: 81
- 被引次数: 0

