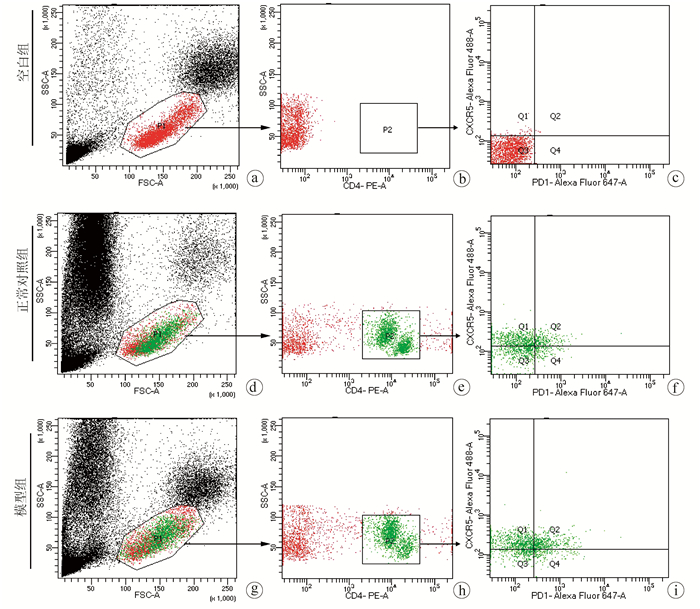
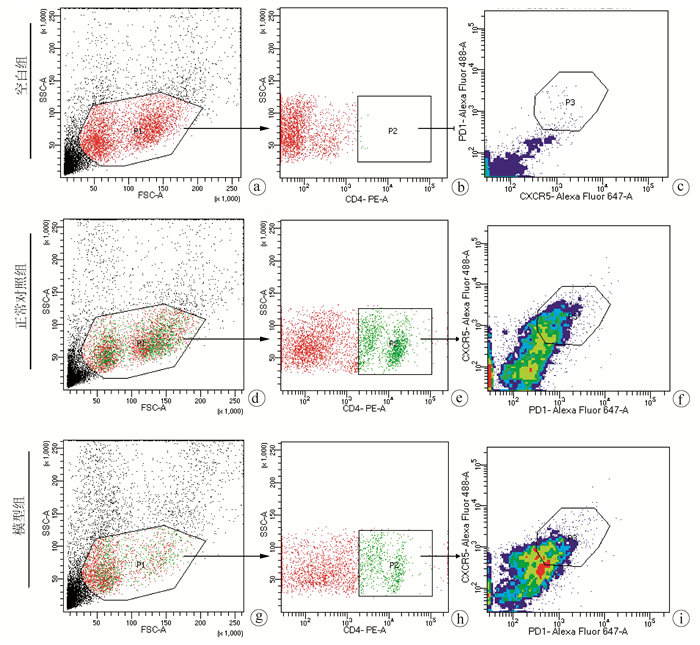
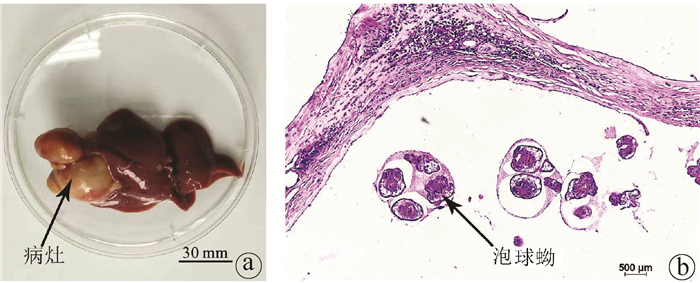
Objective To investigate the level of follicular helper T (Tfh) cells in the peripheral blood and spleen of the host after Echinococcus multilocularis infection and its association with the progression of echinococcosis.Methods A total of 20 Sprague-Dawley rats were randomly divided into normal control group and model group, with 10 rats in each group. The rats in the model group were inoculated with about 2000 protoscoleces in the right liver under direct-view laparotomy, and those in the control group were not given any treatment. The rats were anesthetized and sacrificed after 3 months to collect peripheral blood and spleen cells, and with CD4+CXCR5+PD1+ as the marker of Tfh cells, flow cytometry was used to measure the level of Tfh cells in peripheral blood and spleen. The t-test was used for comparison of Tfh cells between the two groups.Results After 3 months of Echinococcus multilocularis infection, marked lesions were observed in the liver, and HE staining showed the presence of protoscoleces in the lesions. The proportion of CD4+CXCR5+PD1+Tfh cells in CD4+ cells in peripheral blood was 25.63%±3.47% in the model group and 11.12%±2.94% in the normal control group (t=10.230, P < 0.001), a nd the model group had a significantly lower proportion of CD4+CXCR5+PD1+Tfh cells in all cells than the normal control group (0.08%±0.02% vs 0.18%±0.05%, t=5.520, P < 0.001). For the model group, the proportion of CD4+CXCR5+PD1+Tfh cells in all cells in the spleen decreased to 3.00%±0.42%, which was significantly lower than the proportion of 5.30%±1.40% in the normal control group (t=4.769, P < 0.001).Conclusion Tfh cells in peripheral blood are closely associated with the progression of echinococcosis and are expected to become an indicator of Echinococcus multilocularis infection.
CASULLI A, BARTH T, TAMAROZZI F. Echinococcus multilocularis[J]. Trends Parasitol, 2019, 35(9): 738-739. DOI: 10.1016/j.pt.2019.05.005.
[2]
WEN H, VUITTON L, TUXUN T, et al. Echinococcosis: Advances in the 21st Century[J]. Clin Microbiol Rev, 2019, 32(2): e00075-18. DOI: 10.1128/CMR.00075-18.
[3]
CAI H, GUAN Y, MA X, et al. Epidemiology of echinococcosis among schoolchildren in golog tibetan autonomous prefecture, qinghai, China[J]. Am J Trop Med Hyg, 2017, 96(3): 674-679. DOI: 10.4269/ajtmh.16-0479.
[4]
WANG J, GOTTSTEIN B. Immunoregulation in larval Echinococcus multilocularis infection[J]. Parasite Immunol, 2016, 38(3): 182-192. DOI: 10.1111/pim.12292.
[5]
GLATMAN ZARETSKY A, TAYLOR JJ, KING IL, et al. T follicular helper cells differentiate from Th2 cells in response to helminth antigens[J]. J Exp Med, 2009, 206(5): 991-999. DOI: 10.1084/jem.20090303.
[6]
HE L, GU W, WANG M, et al. Extracellular matrix protein 1 promotes follicular helper T cell differentiation and antibody production[J]. Proc Natl Acad Sci U S A, 2018, 115(34): 8621-8626. DOI: 10.1073/pnas.1801196115.
[7]
CROTTY S. T follicular helper cell biology: A decade of discovery and diseases[J]. Immunity, 2019, 50(5): 1132-1148. DOI: 10.1016/j.immuni.2019.04.011.
[8]
CROTTY S. T follicular helper cell differentiation, function, and roles in disease[J]. Immunity, 2014, 41(4): 529-542. DOI: 10.1016/j.immuni.2014.10.004.
[9]
SONG W, CRAFT J. T follicular helper cell heterogeneity: Time, space, and function[J]. Immunol Rev, 2019, 288(1): 85-96. DOI: 10.1111/imr.12740.
[10]
VINUESA CG, LINTERMAN MA, YU D, et al. Follicular helper T cells[J]. Annu Rev Immunol, 2016, 34: 335-368. DOI: 10.1146/annurev-immunol-041015-055605.
[11]
HELMOLD HAIT S, VARGAS-INCHAUSTEGUI DA, MUSICH T, et al. Early T follicular helper cell responses and germinal center reactions are associated with viremia control in immunized rhesus macaques[J]. J Virol, 2019, 93(4). DOI: 10.1128/JVI.01687-18.
[12]
XU W, ZHAO X, WANG X, et al. The Transcription factor tox2 drives T follicular helper cell development via regulating chromatin accessibility[J]. Immunity, 2019, 51(5): 826-839.e5. DOI: 10.1016/j.immuni.2019.10.006.
[13]
WANG B, LI H, SA RL, et al. The expression of ICOS in Tfh cells and the effect of ICOS blocker on the expression of IL-21 in Tfh cells with liver fibrosis[J]. Int J Immunol, 2020, 43(5): 483-487. DOI: 10.3760/cma.j.issn.1673-4394.2020.05.001.
LINDQVIST M, van LUNZEN J, SOGHOIAN DZ, et al. Expansion of HIV-specific T follicular helper cells in chronic HIV infection[J]. J Clin Invest, 2012, 122(9): 3271-3280. DOI: 10.1172/JCI64314.
[15]
VELU V, MYLVAGANAM G, IBEGBU C, et al. Tfh1 cells in germinal centers during chronic HIV/SIV infection[J]. Front Immunol, 2018, 9: 1272. DOI: 10.3389/fimmu.2018.01272.
[16]
GLATMAN ZA, TAYLOR JJ, KING IL, et al. T follicular helper cells differentiate from Th2 cells in response to helminth antigens[J]. J Exp Med, 2009, 206(5): 991-999. DOI: 10.1126/sciimmunol.aan8884.
[17]
KLEIN F, MOUQUET H, DOSENOVIC P, et al. Antibodies in HIV-1 vaccine development and therapy[J]. Science, 2013, 341(6151): 1199-1204. DOI: 10.1126/science.1241144.
[18]
HANSEN DS, OBENG-ADJEI N, LY A, et al. Emerging concepts in T follicular helper cell responses to malaria[J]. Int J Parasitol, 2017, 47(2-3): 105-110. DOI: 10.1016/j.ijpara.2016.09.004.
[19]
DÍAZ A, CASARAVILLA C, ALLEN JE, et al. Understanding the laminated layer of larval Echinococcus Ⅱ: Immunology[J]. Trends Parasitol, 2011, 27(6): 264-273. DOI: 10.1016/j.pt.2011.01.008.
[20]
HOU YJ, ZHANG LQ, FAN HN. Research advances in circulating free DNA in liver cancer and liver-related parasitic diseases[J]. J Clin Hepatol, 2020, 36(2): 430-432. DOI: 10.3969/j.issn.1001-5256.2020.02.043.
WANG ZX, GOU P, YU WH, et al. Measurement and bioinformatics analysis of exosomes microRNAs in bile of hepatic alveolar echinococcosis patients with biliary tract invasion[J]. J Clin Hepatol, 2020, 36(9): 2045-2049. DOI: 10.3969/j.issn.1001-5256.2020.09.027.
DING JB, LI YJ, ZHANG FB. Research progress of hydatidosis immunity and vaccine[J]. J Xinjiang Med Univ, 2019, 42(1): 24-28. DOI: 10.3969/j.issn.1009-5551.2019.01.005.
CAI X, CHEN JJ, TANG DL, et al. Value of alpha-fetoprotein combined with gamma-glutamyl transpeptidase/aspartate aminotransferase ratio in diagnosis of HBV-associated hepatocellular carcinoma[J]. J Clin Hepatol, 2021, 37(9): 2109-2112. DOI: 10.3969/j.issn.1001-5256.2021.09.021.
CAI X, CHEN JJ, TANG DL, et al. Value of alpha-fetoprotein combined with gamma-glutamyl transpeptidase/aspartate aminotransferase ratio in diagnosis of HBV-associated hepatocellular carcinoma[J]. J Clin Hepatol, 2021, 37(9): 2109-2112. DOI: 10.3969/j.issn.1001-5256.2021.09.021.

 下载:
下载:







 DownLoad:
DownLoad: