Value of single or combined measurement of serum PCT,peripheral WBC,and WBC/PCT for early diagnosis of bacterial infection in patients with liver cirrhosis
Objective To investigate the early diagnostic value of single or combined measurement of serum procalcitonin( PCT),peripheral white blood cells( WBC),and ratio of WBC to platelets( PLT)( WBC / PLT) for spontaneous bacterial peritonitis( SBP) in patients with liver cirrhosis. Methods The clinical data of 129 patients with liver cirrhosis who were admitted to our hospital from January 2011 to June2013 were retrospectively analyzed. One hundred and twelve patients who died of liver cirrhosis were divided into infection group,in which the cause of death was complicated infection( n = 94,including 61 SBP cases),and non- infection group( n = 18). Seventeen patients with compensated cirrhosis were assigned to control group. Before treatment with antibiotics,routine bacterial culture was made,and serum PCT,WBC,and WBC / PLT were measured. Mean comparison was made by t test and chi- square analysis. Ratio comparison was made by Pearson χ2test. The receiver operating characteristic( ROC) curve was plotted to calculate the sensitivity and specificity. Results There were 66 patients in the infection group whose bacterial culture was positive,and the positive rates for cultures from blood,ascites and other exudates were 25. 8%,30. 3%,and 43. 9%,respectively. In the infection group,lung infection,SBP,and unknown focus of infection accounted for 8. 5%,64. 9%,and 26. 6% of total cases,respectively. The level of serum PCT in the infection group was significantly higher than those in the non- infection group and the control group( F = 10. 98,P < 0. 05),but showed no significant difference in patients with different sites of infection. When PCT was ≥0. 5 ng / ml,the sensitivity and specificity for diagnosis of complicated infection were 92. 5%and 77. 1%,respectively,and the area under the ROC curve was 0. 89. When PCT was ≥2 ng / ml,the sensitivity was 62. 7% and the specificity was 94. 2%. When the peripheral WBC count was ≥10 × 109/ L and the WBC / PLT was ≥0. 25,the sensitivity was 47. 8% and39. 6%,respectively,and the specificity was 100%. When PCT was combined with WBC / PLT for diagnosis of complicated infection,the sensitivity was 76. 8% and the specificity was 94. 2%. For patients with SBP,the levels of PCT and WBC and WBC / PLT showed no significant differences between positive and negative ascites cultures. When PCT was combined with WBC / PLT for diagnosis of SBP,the sensitivity was 83. 6% and the specificity was 94. 2%. Conclusion PCT and WBC / PLT can be used as early diagnostic indicators for complicated infection in cirrhotic patients. PCT combined with WBC / PLT has a significant value for early diagnosis of SBP in cirrhotic patients.
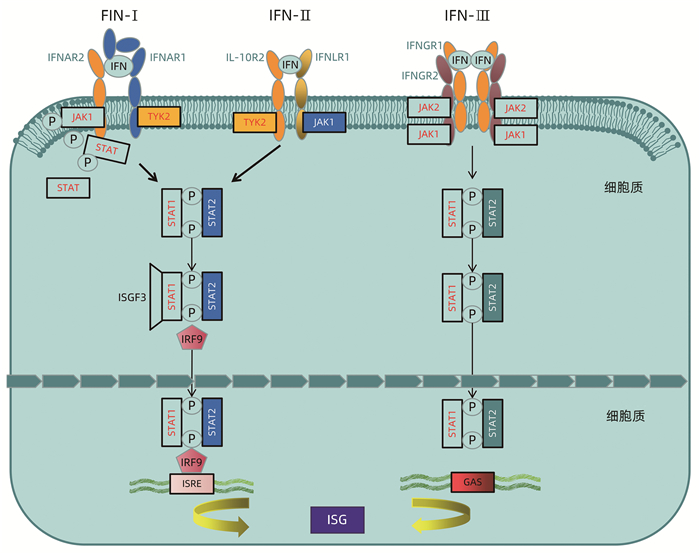
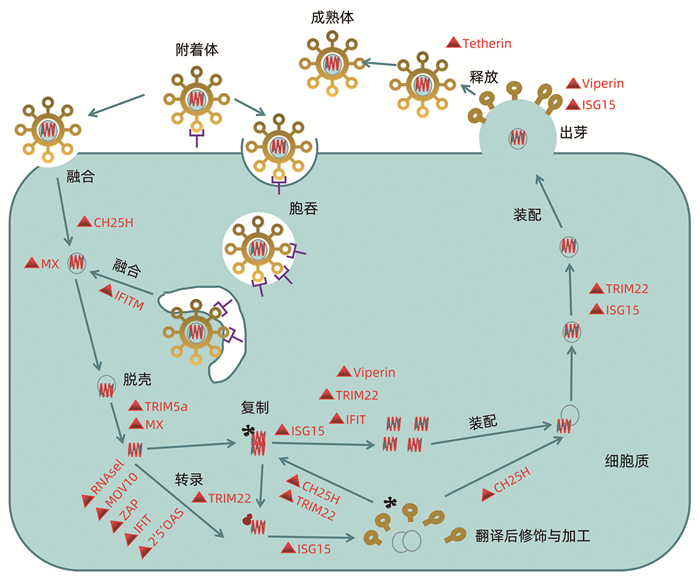
随着对病原体相关分子模式(pathogen-associated molecular patterns, PAMP)和模式识别受体(pattern recognition receptors, PRRS)的认识,发现病毒侵入细胞后,通过Toll样受体(TLR)和RIG-1样受体(IRG like receptor, RLR)对病毒核酸等病原体的特征分子进行识别,进而激活一系列的下游信号转导分子,最终诱导细胞转录因子的激活或IFN等效应蛋白的产生。机体的其他信号传导蛋白如PKR、维甲酸诱导基因1等可被IFN刺激高表达,增强机体对病原识别和检测能力,同时ISG可以增强IFN的免疫应答信号[16]。常见的具有正调控功能的ISG包括OAS,PKR,IRF3、7、9及STAT1/2等。
3.2
负向调控IFN免疫应答
部分ISG可通过抑制IFN介导的下游信号通路负向调控IFN免疫应答。泛素特异性蛋白酶18 (USP18/UBP43)是泛素化特异性蛋白酶家族成员之一,是ISGl5的特异性水解酶,USP18通过竞争性与IFNAR2亚基结合,进而抑制JAK1与IFNAR2结合,进一步抑制IFN-Ⅰ诱导的Jak-STAT信号通路发挥抵抗IFN的抗病毒治疗[28]。细胞因子信号抑制物(suppressor of cytokine signaling,SOCS)可被IFN正向调节表达,但SOCS的过表达又可以抑制IFN通路的Jak的磷酸化及STAT的活化[29]。
Sun Jing, Po JunJie, Fan YuZe, He WeiPing, Li Huan, Sun YingZhe, Wang Hui, Duan XueZhang. Effect of stereotactic body radiotherapy versus intensity-modulated radiotherapy in primary liver cancer patients with secondary malignant tumor of vertebra[J]. J Clin Hepatol, 2016, 32(6): 1135-1138. DOI: 10.3969/j.issn.1001-5256.2016.06.023.
Sun Jing, Po JunJie, Fan YuZe, He WeiPing, Li Huan, Sun YingZhe, Wang Hui, Duan XueZhang. Effect of stereotactic body radiotherapy versus intensity-modulated radiotherapy in primary liver cancer patients with secondary malignant tumor of vertebra[J]. J Clin Hepatol, 2016, 32(6): 1135-1138. DOI: 10.3969/j.issn.1001-5256.2016.06.023.

 下载:
下载:






 DownLoad:
DownLoad: