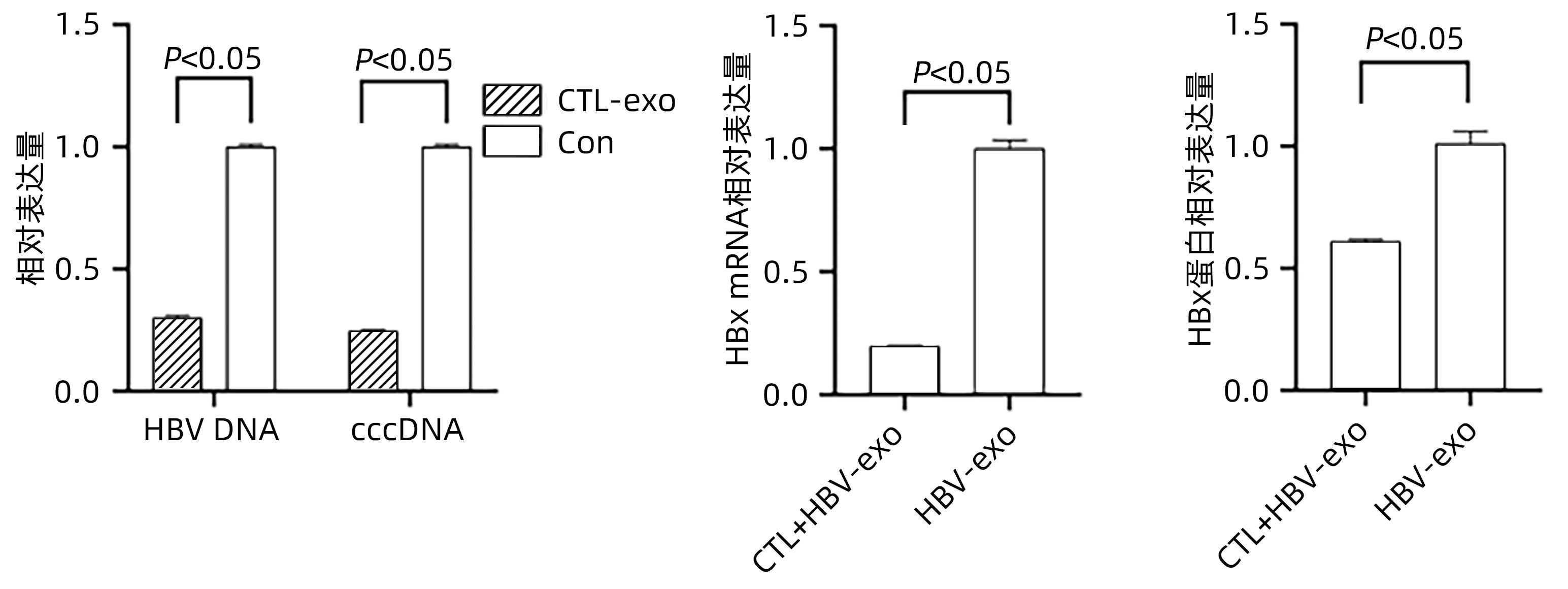
Objective To investigate whether cytotoxic T lymphocyte (CTL)-derived exosomes can downregulate HBx expression and inhibit hepatic stellate cell (HSC) activation.Methods The supernatants of HepG2, HepGA14, and CTL cells were collected to extract exosomes, which were referred to as NC-exo, HBV-exo, and CTL-exo, respectively). Transmission electron microscopy was used to observe their morphology, and Western Blot was used to measure the expression of the markers of exosomes CD63 and TSG101. NC-exo, HBV-exo, and CTL-exo labeled by BODIPY dye were mixed with HBV-exo at different ratios and were then co-cultured with HSC LX-2 (HSC-LX2). A fluorescence microscope was used to observe whether exosomes could enter LX-2 cells, and an fluorescence microscope was used to observe cell morphological changes; quantitative real-time PCR (qPCR) was used to measure the expression of the activated biomarkers such as transforming growth factor-β1 (TGF-β1), ɑ-smooth muscle actin (ɑ-SMA), and collagen type I (Collagen I) in LX-2 cells. CTL-exo was added to the HepGA14 culture system; then qPCR was used to measure the mRNA expression level of HBV DNA, cccDNA, and HBx in exosomes in HepGA14 cells, and Western Blot was used to measure the protein expression level of HBx in exosomes. The t-test was used for comparison of normally distributed continuous data between two groups; a one-way analysis of variance was used for comparison between multiple groups, and the least significant difference t-test was used for further comparison between two groups.Results The exosomes were all microcysts with a double-layer membrane structure and were circular or elliptical in shape, with the expression of the signature proteins CD63 and TSG101, and the vesicles had a diameter of 50-100 nm. The fluorescence microscope showed that exosomes could enter LX-2 cells, and HSC were enlarged with extended cell processes. The results of qPCR showed that there were significant differences in the expression levels of TGF-β1, ɑ-SMA, and Collagen I genes between the NC-exo, HBV-exo, NC-exo+HBV-exo, and Con groups (F=444.678, 417.144, and 571.508, all P<0.05). After the intervention of HepGA14 cells with CTL-exo, qPCR results showed that compared with the control group, there were significant reductions in the expression levels of HBV DNA and cccDNA in HepGA14 cells (all P<0.05), the relative mRNA expression level of HBx in exosomes (P<0.05), and the protein expression level of HBx (P<0.05). CTL-exo and HBV-exo were mixed at different ratios (2∶1, 5∶1, 10∶1) and were then used for the intervention of LX-2 cells, and qPCR results showed that the expression levels of TGF-β1, ɑ-SMA, and Collagen I genes in LX-2 cells gradually decreased with the increase in the ratio of CTL-exo between groups (P<0.05).Conclusion CTL-exo can downregulate the protein expression of HBx in HBV-exo to inhibit HSC activation, suggesting that CTL-exo has an anti-hepatitis B liver fibrosis effect.
LI T, YAN Y, WANG B, et al. Exosomes derived from human umbilical cord mesenchymal stem cells alleviate liver fibrosis[J]. Stem Cells Dev, 2013, 22( 6): 845- 854. DOI: 10.1089/scd.2012.0395.
[2]
Chinese Society of Infectious Diseases, Chinese Medical Association, Chinese Society of Hepatology, Chinese Medical Association. Guidelines for the prevention and treatment of chronic hepatitis B(version 2019)[J]. J Clin Hepatol, 2019, 35( 12): 2648- 2669. DOI: 10.3969/j.issn.1001-5256.2019.12.007.
LIU J, LIANG W, JING W, et al. Countdown to 2030: eliminating hepatitis B disease, China[J]. Bull World Health Organ, 2019, 97( 3): 230- 238. DOI: 10.2471/BLT.18.219469.
[4]
LIU JW, XIE J, LI TS. Specific T-lymphocyte response and clinical outcome of patients with hepatitis B virus infection[J]. Med J Peking Union Med Coll Hosp, 2012, 3( 3): 350- 353. DOI: 10.3969/j.issn.1674-9081.2012.03.021.
MA S, CHEN X, TAN Q, et al. An engineered novel lentivector specifically transducing dendritic cells and eliciting robust HBV-specific CTL response by upregulating autophagy in T cells[J]. Cell Cycle, 2018, 17( 10): 1220- 1234. DOI: 10.1080/15384101.2018.1471312.
[6]
CHENG ST, REN JH, CAI XF, et al. HBx-elevated SIRT2 promotes HBV replication and hepatocarcinogenesis[J]. Biochem Biophys Res Commun, 2018, 496( 3): 904- 910. DOI: 10.1016/j.bbrc.2018.01.127.
[7]
CHEN HY, CHEN ZX, HUANG RF, et al. Hepatitis B virus X protein activates human hepatic stellate cells through upregulating TGFβ1[J]. Genet Mol Res, 2014, 13( 4): 8645- 8656. DOI: 10.4238/2014.October.27.4.
[8]
QIU H, LIN DY, LI JY. Screening and identification of dominant monoclonal HepG2 cell strain with 1.3-fold HBV genome[J]. World Chin J Dig, 2021, 29( 16): 934- 944. DOI: 10.11569/wcjd.v29.i16.934.
ROCKEY DC. Liver fibrosis reversion after suppression of hepatitis B virus[J]. Clin Liver Dis, 2016, 20( 4): 667- 679. DOI: 10.1016/j.cld.2016.06.003.
[11]
CHANG TT, LIAW YF, WU SS, et al. Long-term entecavir therapy results in the reversal of fibrosis/cirrhosis and continued histological improvement in patients with chronic hepatitis B[J]. Hepatology, 2010, 52( 3): 886- 893. DOI: 10.1002/hep.23785.
[12]
TZIOMALOS K. Effect of antiviral treatment on the risk of hepatocellular carcinoma in patients with chronic hepatitis B[J]. World J Hepatol, 2010, 2( 3): 91- 93. DOI: 10.4254/wjh.v2.i3.91.
[13]
CHEN BW, ZHU XJ, ZHANG X, et al. Influence of virologic response on disease progression in patients with compensated hepatitis B cirrhosis[J]. J Clin Hepatol, 2021, 37( 8): 1811- 1816. DOI: 10.3969/j.issn.1001-5256.2021.08.014.
GIERSCH K, ALLWEISS L, VOLZ T, et al. Serum HBV pgRNA as a clinical marker for cccDNA activity[J]. J Hepatol, 2017, 66( 2): 460- 462. DOI: 10.1016/j.jhep.2016.09.028.
[15]
MOHAMMADI A, TAJIK N, SHAH-HOSSEINI A, et al. FAS and FAS-ligand promoter polymorphisms in hepatitis B virus infection[J]. Hepat Mon, 2015, 15( 10): e26490. DOI: 10.5812/hepatmon.26490.
[16]
KONDO Y, KOBAYASHI K, ASABE S, et al. Vigorous response of cytotoxic T lymphocytes associated with systemic activation of CD8 T lymphocytes in fulminant hepatitis B[J]. Liver Int, 2004, 24( 6): 561- 567. DOI: 10.1111/j.1478-3231.2004.0982.x.
[17]
HAQUE M, LEI F, XIONG X, et al. Stem cell-derived viral antigen-specific T cells suppress HBV replication through production of IFN-γ and TNF-α[J]. iScience, 2020, 23( 7): 101333. DOI: 10.1016/j.isci.2020.101333.
[18]
MOON B, CHANG S. Exosome as a delivery vehicle for cancer therapy[J]. Cells, 2022, 11( 3): 316. DOI: 10.3390/cells11030316.
[19]
COCUCCI E, MELDOLESI J. Ectosomes and exosomes: shedding the confusion between extracellular vesicles[J]. Trends Cell Biol, 2015, 25( 6): 364- 372. DOI: 10.1016/j.tcb.2015.01.004.
[20]
JIA Y, LI Y, WANG YJ. Research progress on exosome targeted delivery of nucleic acid molecules[J]. China Med Herald, 2023, 20( 3): 33- 36, 49. DOI: 10.20047/j.issn1673-7210.2023.03.07.
SUN JS, LU MY. Research progress on the formation of exosomes and their rolesin immune regulation[J]. J Tianjin Normal University(Natural Science Edition), 2020, 40( 1): 9- 15. DOI: 10.19638/j.issn1671-1114.20200102.
ABDULKARIM AS, CAO H, HUANG B, et al. The large GTPase dynamin is required for hepatitis B virus protein secretion from hepatocytes[J]. J Hepatol, 2003, 38( 1): 76- 83. DOI: 10.1016/s0168-8278(02)00326-4.
WANG J, WANG YC, WANG CY. Association between monocyte-to-high-density lipoprotein cholesterol ratio and nonalcoholic fatty liver disease[J]. J Clin Hepatol, 2021, 37(5): 1137-1141. DOI: 10.3969/j.issn.1001-5256.2021.05.031.
WANG J, WANG YC, WANG CY. Association between monocyte-to-high-density lipoprotein cholesterol ratio and nonalcoholic fatty liver disease[J]. J Clin Hepatol, 2021, 37(5): 1137-1141. DOI: 10.3969/j.issn.1001-5256.2021.05.031.

 下载:
下载:







 DownLoad:
DownLoad: