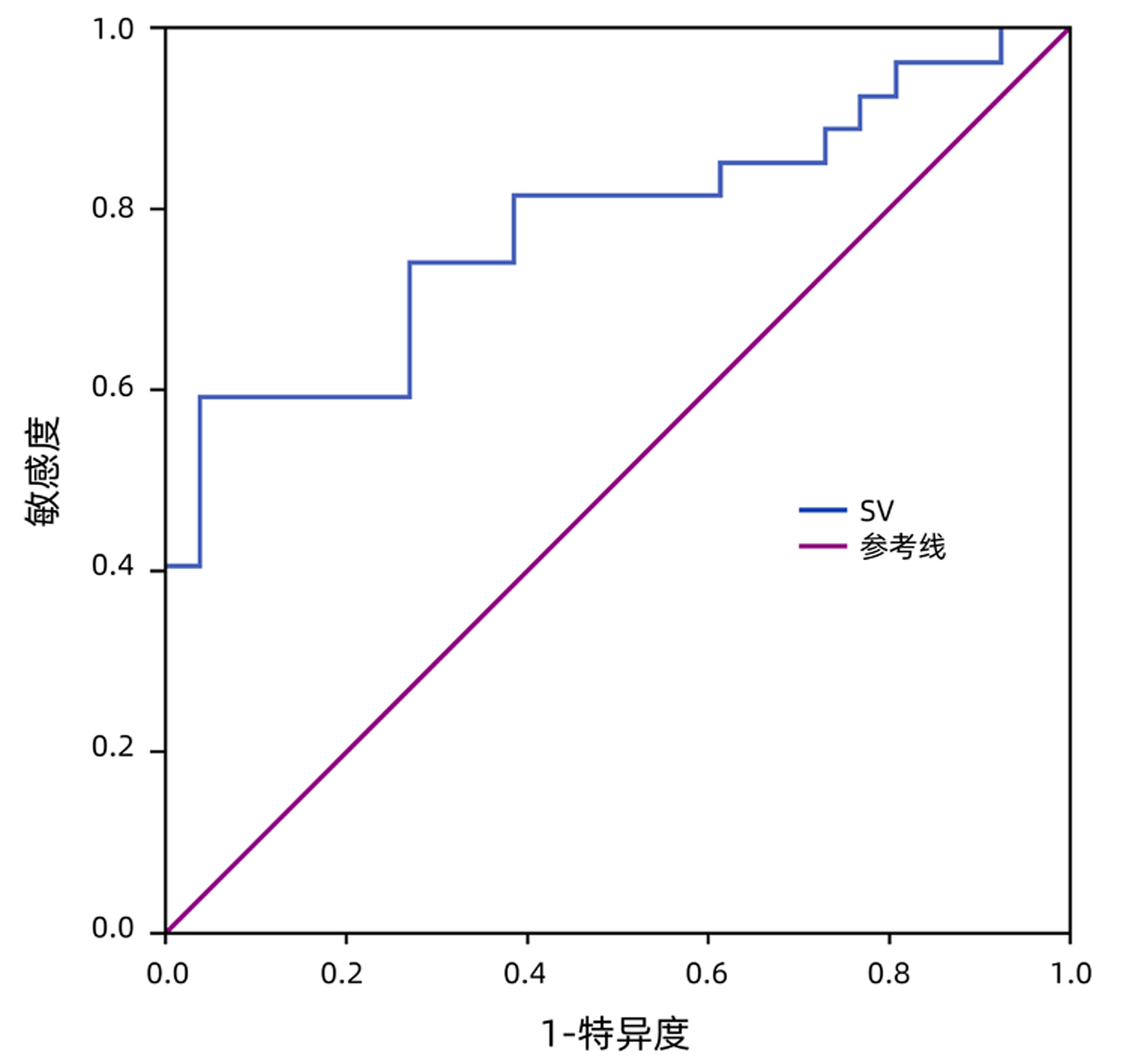
| [1] |
|
| [2] |
ROCKEY DC. An update: Portal hypertensive gastropathy and colopathy[J]. Clin Liver Dis, 2019, 23(4): 643-658. DOI: 10.1016/j.cld.2019.07.002. |
| [3] |
LYLES T, ELLIOTT A, ROCKEY DC. A risk scoring system to predict in-hospital mortality in patients with cirrhosis presenting with upper gastrointestinal bleeding[J]. J Clin Gastroenterol, 2014, 48(8): 712-720. DOI: 10.1097/MCG.0000000000000014. |
| [4] |
|
| [5] |
AMER IF, EL SHENNAWY EM, EL BATEA H, et al. Accuracy of noninvasive tests in the prediction of portal hypertensive gastropathy in Egyptian patients with cirrhosis[J]. JGH Open, 2021, 5(2): 286-293. DOI: 10.1002/jgh3.12486. |
| [6] |
EL-KALLA F, MANSOUR L, KOBTAN A, et al. Blood ammonia level correlates with severity of cirrhotic portal hypertensive gastropathy[J]. Gastroenterol Res Pract, 2018, 2018: 9067583. DOI: 10.1155/2018/9067583. |
| [7] |
NISHINO K, KAWANAKA M, MANABE N, et al. Portal hypertensive gastropathy in liver cirrhosis: Prevalence, natural history, and risk factors[J]. Intern Med, 2022, 61(5): 605-613. DOI: 10.2169/internalmedicine.7943-21. |
| [8] |
MIN YW, BAE SY, GWAK GY, et al. A clinical predictor of varices and portal hypertensive gastropathy in patients with chronic liver disease[J]. Clin Mol Hepatol, 2012, 18(2): 178-184. DOI: 10.3350/cmh.2012.18.2.178. |
| [9] |
YU S, CHEN W, JIANG Z. Platelet count/spleen volume ratio has a good predictive value for esophageal varices in patients with hepatitis B liver cirrhosis[J]. PLoS One, 2021, 16(12): e0260774. DOI: 10.1371/journal.pone.0260774. |
| [10] |
Chinese Society of Hepatology, Chinese Medical Association. Chinese guidelines on the management of liver cirrhosis[J]. J Clin Hepatol, 2019, 35(11): 2408-2425. DOI: 10.3969/j.issn.1001-5256.2019.11.006. |
| [11] |
DE FRANCHIS R. Updating consensus in portal hypertension: report of the Baveno Ⅲ Consensus Workshop on definitions, methodology and therapeutic strategies in portal hypertension[J]. J Hepatol, 2000, 33(5): 846-852. DOI: 10.1016/s0168-8278(00)80320-7. |
| [12] |
BADRAN DH, KALBOUNEH HM, AL-HADIDI MT, et al. Ultrasonographic assessment of splenic volume and its correlation with body parameters in a Jordanian population[J]. Saudi Med J, 2015, 36(8): 967-972. DOI: 10.15537/smj.2015.8.11809. |
| [13] |
MOROZOV SV, IZRANOV VA. Methods of ultrasound spleen morphometry[J]. J Ultrasound Med, 2022, 41(9): 2123-2133. DOI: 10.1002/jum.15901. |
| [14] |
LI Y, WANG ZH, LI PP, et al. 3D visualization technology for measuring the spleen volume of normal Chinese adult[J]. J Hepatobiliary Surg, 2018, 26(5): 340-343. DOI: 10.3969/j.issn.1006-4761.2018.05.007. |
| [15] |
DE FRANCHIS R. Evolving consensus in portal hypertension. Report of the Baveno Ⅳ consensus workshop on methodology of diagnosis and therapy in portal hypertension[J]. J Hepatol, 2005, 43(1): 167-176. DOI: 10.1016/j.jhep.2005.05.009. |
| [16] |
CHEN SY, LIN QY, HUANG XQ. Studies on diagnosis and pathogenesis of portal hypertensive gastropathy[J]. J Diagn Concepts Pract, 2016, 15(5): 451-454. DOI: 10.16150/j.1671-2870.2016.05.003. |
| [17] |
DE FRANCHIS R, BOSCH J, GARCIA-TSAO G, et al. Baveno Ⅶ-Renewing consensus in portal hypertension[J]. J Hepatol, 2022, 76(4): 959-974. DOI: 10.1016/j.jhep.2021.12.022. |
| [18] |
SALEEM K, BAIG FA, NIDA M, et al. Correlation between severity of portal hypertensive gastropathy and size of oesophageal varices in cirrhotic hepatitis-C patients[J]. J Ayub Med Coll Abbottabad, 2018, 30(1): 54-57.
|
| [19] |
SIMBRUNNER B, BEER A, WÖRAN K, et al. Portal hypertensive gastropathy is associated with iron deficiency anemia[J]. Wien Klin Wochenschr, 2020, 132(1-2): 1-11. DOI: 10.1007/s00508-019-01593-w. |
| [20] |
SUN DY. Prediction of FIB-4、Lok and King score measurement for esophageal varices in cirrhosis[D]. Hefei: Anhui Medical University, 2021.
孙迪一. FIB-4、Lok和King评分对肝硬化食管静脉曲张的预测价值[D]. 合肥: 安徽医科大学, 2021.
|
| [21] |
MANDHWANI R, HANIF FM, UL HAQUE MM, et al. Noninvasive clinical predictors of portal hypertensive gastropathy in patients with liver cirrhosis[J]. J Transl Int Med, 2017, 5(3): 169-173. DOI: 10.1515/jtim-2017-0025. |
| [22] |
JIANG A, LI ZF. Common causes and treatment strategies of hyperslenism[J/OL]. Chin J Hepat Surg (Electronic Edition), 2018, 7(2): 97-99. DOI: 10.3877/cma.j.issn.2095-3232.2018.02.003.
蒋安, 李宗芳. 脾功能亢进常见原因及治疗策略[J/OL]. 中华肝脏外科手术学电子杂志, 2018, 7(2): 97-99. DOI: 10.3877/cma.j.issn.2095-3232.2018.02.003.
|




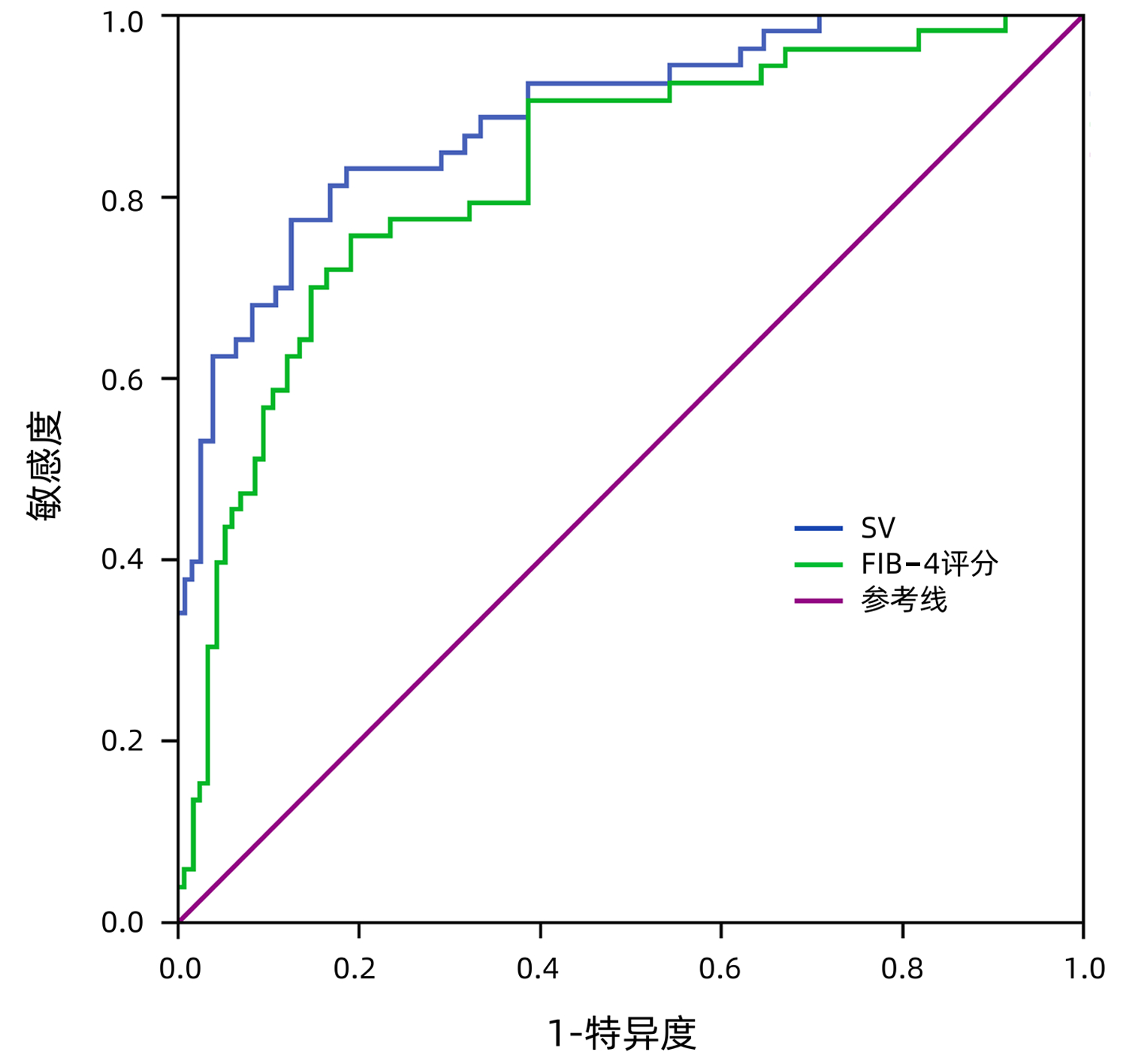




 DownLoad:
DownLoad: